INTRODUCTION
Traumatic brain injury (TBI) is a significant cause of morbidity and mortality in pediatric and adult populations [1,2]. In the United States, TBI affects more than 1.5 million people annually; approximately 300,000 are hospitalized and more than 50,000 die with TBI as a contributing cause, accounting for approximately one-third of injury-related deaths [3,4]. Approximately 475,000 children aged 0 to 14 years are diagnosed with TBI annually in the United States and TBI is the current leading cause of death among children older than 1 year [5].
Regardless of severity, several patients with TBI report some degree of neurobehavioral sequelae after injury [6]. Significant changes in cognitive, behavioral, and emotional functioning are prevalent in patients with TBI. Cognitive complaints are typically reported in the areas of memory, attention, and executive functioning [7]. Additionally, increased irritability, depression, or anxiety is often reported by individuals with TBI and other behavioral changes may include impulsive behaviors, reduced frustration tolerance, lack of empathy, emotional lability, apathy, or aggression [7]. In particular, severe and moderate TBI frequently produces many types of temporary or permanent physical, cognitive, behavioral, and emotional impairments [8]. Initial and persistent cognitive deficits after TBI are the most common complaints, including loss of short-term memory and learning impairment. Furthermore, severe and moderate TBI results in an increased relative risk of various psychiatric disorders, including mood and anxiety disorders [9]. The lack of identification of neurobehavioral sequelae following TBI can lead to no or inappropriate treatment.
Unlike in adults, the pediatric brain undergoes rapid development. Therefore, pediatric neurotrauma is a frequent cause of neurobehavioral morbidity and can result in unique challenges as the consequences of injuries are overlaid on normal developmental changes [10]. In addition, injury mechanisms in the pediatric population differ from those in adults, potentially resulting in unique primary and secondary lesions in the developing brain [11]. Although many factors affect the clinical outcome of patients following TBI, patient age has been found to be one of the most reliable and consistent prognostic factors following TBI in various clinical studies, where older age is associated with worse outcomes [1,12-15]. However, clinical and experimental studies of the prognostic differences between the developing and adult brain following TBI are lacking. Knowledge of the age-specific post-injury neurobehavioral sequelae is essential for understanding and managing children with neurotrauma.
In a recent study, of age-dependent neuronal sensitivities to ischemic injury adult (8 to 12 weeks) and pediatric (postnatal 20 to 25 days) mice were subjected to 6-minute cardiac arrest and cardiopulmonary resuscitation; cardiac arrest produced global cerebral ischemia and neuronal injury in vulnerable brain regions in juvenile mice [16]. Namely, the injured area and degree of injury was different from that in the adult brain. Similarly, the current study hypothesized that TBI has age-dependent neuronal sensitivity to traumatic injury. Therefore, an experimental approach to control various confounding factors is required to investigate the differences in vulnerability to TBI between the pediatric and adult brain.
Different experimental TBI models have been used to reflect the heterogeneous clinical situation in studies of TBI outcomes. Among TBI animal models, the controlled cortical impact (CCI) model can produce a more precise injury by controlling deformation parameters, such as time, velocity, and depth of impact, and thus, has advantages over other TBI models with respect to the absence of a risk for rebound injury, higher accuracy, reproducibility, and simplicity [17]. There are limited published studies describing the differences in neurobehavioral outcomes between pediatric and adult patients after the same magnitude of TBI.
This study assessed the differences in exploratory, cognitive, and depressive-like behaviors between pediatric and adult male mice exposed to moderate to severe CCI. Using neurobehavioral assessments evaluating outcomes after TBI, we expect to (1) understand the effects of injury on cognitive skills, behavior, and learning; and (2) determine how these outcomes are related to the brain insult and any ensuing abnormalities in neural development.
METHODS
Animal population
All experimental protocols were approved by the animal care and use committee of Chungbuk National University, Cheongju, Korea (CBNUR-854-15). We used 8- to 12-week-old adult and 21- to 25-day-old pediatric C57Bl/6 male mice in this study. Mice were housed with a standard 12-hour light:dark cycle and had ad libitum access to food and water.
Surgical procedures
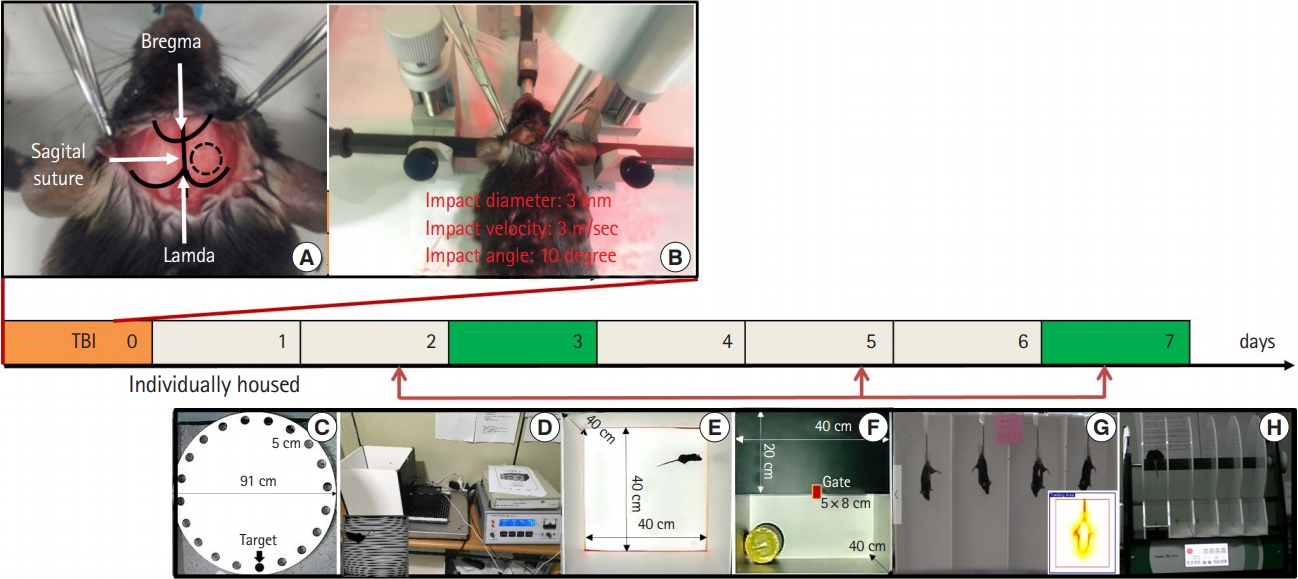
Mice were anesthetized with an intramuscular injection of 15 mg/kg of tiletamine/zolazepam (Zoletil; Virbac, Carros, France). The head of the mouse was shaved between the ears and cleaned using betadine, a midline scalp incision was made, and the right parietal bone was exposed. Next, a 4-mm diameter circle was drawn centered between the lambda and bregma 0.5 mm right of the midline. A right parietal craniotomy was carefully performed using a surgical microscope and a micromotor drill (Stoelting, Wood Dale, IL, USA) along the marked circle. Next, the CCI device was calibrated with respect to the exposed dura mater within the craniotomy. The parameters of the impact applied to the experimental animals were a depth of 2.0 mm, mean velocity of 3.0 m/sec, and duration of 500 ms. After the impact, the scalp incision was sutured with 5-0 nylon. The sham-operated groups received a craniotomy without CCI injury. The surgical procedure is shown in Fig. 1.
Health assessment score
Mice were weighed daily and a health assessment score was recorded for each mouse after surgery. The graded scoring systems ranged from 0 to 2, 0 to 3, or 0 to 5 depending on the behavior assessed, with 0 indicating no deficit and higher values indicating significant impairment. The behaviors assessed included consciousness (0ŌĆō3), interaction (0ŌĆō2), ability to grab a wire top (0ŌĆō2), motor function (0ŌĆō5), and activity (0ŌĆō2) [18,19]. Scores in each category were summated to generate an overall health assessment score.
Hematoxylin and eosin staining and estimations of cortical lesion volume
At 3 and 7 days after TBI, animals were anesthetized with 15 mg/kg of tiletamine/zolazepam and transcardially perfused with cold 0.9% saline followed by 10% formalin. Brains were removed, post-fixed with 10% formalin, and embedded in paraffin. Coronal sections, 6-╬╝m thick, were collected at 100-micron intervals and stained with hematoxylin and eosin (H&E). Micrographs of the H&E-stained sections were obtained to record and calculate the extent of injury at a magnification of 1.25├Ś using a CX43 Olympus microscope (Olympus, Tokyo, Japan).
Cortical lesion volumes (mm3) were determined by calculating the area of the lesion (mm2) at a magnification of ├Ś1.25 using a CX43 microscope. This was done by outlining the missing cortical tissue for each section sliced at 500-╬╝m intervals, and then multiplying the sum of lesion areas obtained from each section with the distance between sections. Lesion volumes were analyzed using ImageJ ver. 1.42 (National Institutes of Health, Bethesda, MD, USA).
The hippocampal lesion volume ratio (%) was determined by calculating the relative injury ratio (%) after comparison with the non-injured left hippocampus.
Behavioral testing
Locomotor activity
Locomotor activity was measured for 30 minutes in a white open top acrylic box (40├Ś40├Ś40 cm) with an illumination intensity of 20 lux at the box floor level. The activity was automatically recorded using the video tracking system using SMART 3.0 software (Panlab/Harvard Apparatus, Holliston, MA, USA). The distance moved (mm), time spent in the center (25%), time spent in the area margins, and mean walking speed (cm/sec) were also evaluated.
Barnes maze test
The Barnes maze test was conducted as previously described with minor modifications [20]. The maze consisted of a white, acrylic, circular platform (91-cm diameter) with 20 equally spaced holes and a black acrylic escape box (20├Ś5├Ś6 cm) along the perimeter. The maze was surrounded by four spatial cues at the height of the maze.
Acquisition trials
Each mouse was trained for four acquisition trials per day over 3 days with an inter-trial interval of 10 to 15 minutes. Immediately prior to the first trial, the mouse was placed in the middle of the maze in a black-colored starting cylinder (10-cm diameter) and a buzzer (80 to 90 dB) was turned on. After 10 seconds, the chamber was lifted, and the mouse was pre-trained to enter the escape box by guiding it to the escape box and remaining there for 2 minutes. Following the pre-training trial, the first trial was initiated.
At the beginning of each trial, the mouse was placed in the same starting chamber, and 10 seconds after the onset of a buzzer and light, the chamber was lifted, and the mouse was free to explore the maze. The trial ended when the mouse entered the goal tunnel or after 3 minutes into the trial. Immediately after the mouse entered the tunnel, the buzzer was turned off and the mouse could stay in the tunnel for 1 minute.
After each trial, the entire maze was cleaned with 70% alcohol and rotated to eliminate any intra-maze cues. Trials were recorded using a video tracking system with SMART software 3.0.
Passive-avoidance test
Cognitive function was assessed using a computerized one-trial-learning passive avoidance test (PAT; Panlab/Harvard Apparatus, Barcelona, Spain). The apparatus consisted of a square white compartment (25├Ś25├Ś24 cm) connected to an enclosed dark compartment (13.5├Ś7.5├Ś7.5 cm) by a motorized sliding door. On day 3 after CCI, all mice received acquisition training. They were placed in the bright chamber; after 5 seconds the automatic door between the two chambers was opened. Once the mice entered the dark chamber, the door was closed and the mouse received a 1 mA foot shock for 2 seconds via the grid floor. After delivery of the foot shock, the mice were left in the dark compartment for another 10 seconds. One day or 5 days after acquisition training, the mice were tested for retention of the task. They were placed in the bright chamber of the apparatus, and after 5 seconds, the door was opened. Latency to enter the dark chamber was recorded to a maximum of 300 seconds. Animals that did not cross into the dark chamber during the testing period were assigned a latency of 300 seconds.
Light/dark test
The apparatus consisted of a black compartment (20├Ś40├Ś40 cm) and a white compartment (20├Ś40├Ś40 cm) separated by a connecting gate (5├Ś8 cm). Each animal was individually placed at the center of the bright compartment (facing away from the door) and the following parameters were measured for 5 minutes: latency of the initial movement from the light to dark area (latency of transition), total number of transitions between light and dark areas, total time spent in the light area.
Tail suspension test
One week after CCI, the mice were subjected to the tail suspension test, performed as previously described [21]. Briefly, mice were individually suspended by the tail to a hook using adhesive tape and recorded by a video camera placed directly in front of the mice for subsequent analysis. The measured behavioral score was immobility during the entire 6-minute test session, using the video tracking system with the SMART software 3.0.
Rotarod testing
Animals were tested on a 7-cm diameter Rotarod treadmill (LE 8500; Panlab SL, Barcelona, Spain) elevated 50 cm above the bottom of the apparatus and attached to a motor to control speed. Briefly, mice were placed on the rotarod at a starting speed of 4 rpm/min and acceleration of 0.5 rpm/sec; the mice were subjected to three trials. In each trial, animals were scored for their latency to fall (in seconds) and speed on fall (m/sec). Animals rested a minimum of 1 hour between trials to avoid fatigue. The average latency to fall for the three trials was used in analyses.
Statistical analysis
All data are presented as mean┬▒standard error of the mean. Statistical evaluation of the data were performed using two-way analysis of variance (ANOVA) and post hoc TurkeyŌĆÖs test. The Kaplan-Meier method was used for survival analysis. P<0.05 was considered statistically significant. Data were analysed using the PASW/SPSSTM software, version 18 (IBM Inc., Chicago, USA) and GraphPad Prism 5.01 (GraphPad Prism Software, San Diego, CA, USA).
RESULTS
Physiological variables
Measures including body weight, body temperature during surgery, age, survival rates, and health assessment score are shown in Table 1. There was no significant difference in body weight, body temperature during surgery, or age between the sham and 2-mm CCI for both adult and pediatric groups. Weight gain (%) in the pediatric group was significantly different from that in the adult group on postoperative day 7. However, there was no significant difference in the health assessment score during postoperative day 1 to 3. Fig. 2 shows that the survival rate after surgery among pediatric mice was lower than that in the adult mice. The difference in survival rate was not surprising because pediatric mice during the weaning period were young & small, and may be more sensitive to surgical complications.
Cortical impact injury volumes
To determine injury severity according to the depth of impact, we first impacted the mouse cortex with three different depths of impact (0.0, 1.0, and 2.0 mm). Analysis of the tissue samples obtained 7 days after injury demonstrated a progressively larger lesion of the brain proportional to the depth of impact. Sham mice showed no or minimal injury due to drilling. All mice with 2-mm depth of impact showed apparent injury to the hippocampus, in marked contrast to mice with 1-mm depth of impact that had an intact hippocampus, as seen on H&E staining (Fig. 3A). Therefore, we used a 2-mm depth of impact (severe and moderate injury) to analyze differences between adult and pediatric groups. The procedure is presented in Fig. 3.
In the measurement of the injury volume of the cortex and hippocampus in adult and pediatric mice, a statistically significant difference was not observed between the adult and pediatric male mice at 3 and 7 days following 2-mm depth of impact CCI (Fig. 3B). However, there was a difference in the injury volume between 3 and 7 days after CCI in the adult groups (Fig. 3B).
Neurobehavioral outcomes
Spatial learning and memory deficits in the Barnes maze
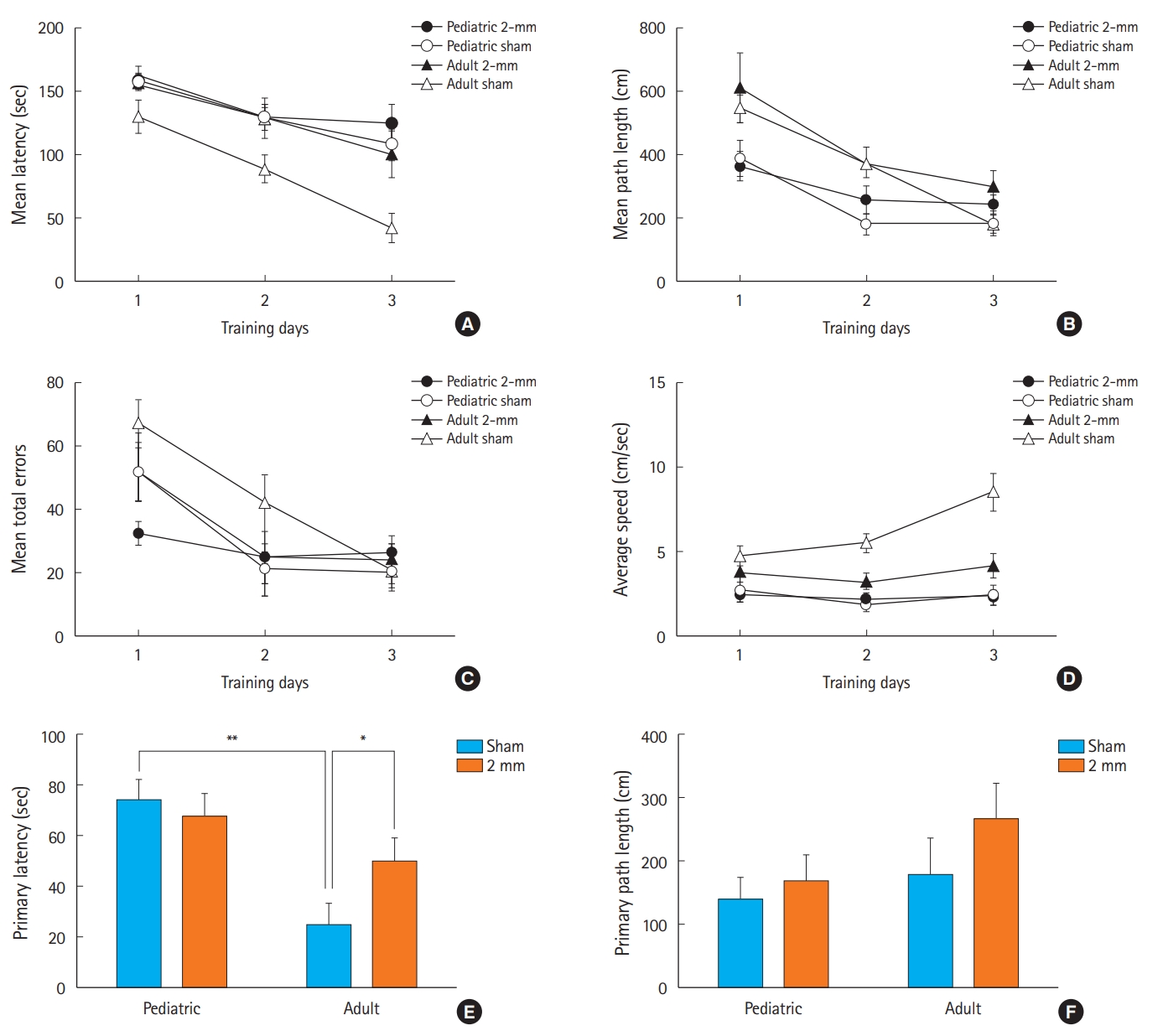
Acquisition phase: During training, there was a significant difference in latency to enter the target hole between sham and 2-mm TBI adult mice. Latency significantly decreased during the three training days, indicating a mean performance improvement in sham adult mice. Interestingly, pediatric mice did not show a significant difference in latency or latency decrease between the sham and 2-mm CCI groups during training (Fig. 4A).
The distance (cm) travelled (Fig. 4B) to enter the target hole and mean total number of errors (Fig. 4C), and average speed (Fig. 4D) were analyzed using a repeated measures ANOVA; these values were found to significantly decrease during the acquisition phase in the sham adult mice but not in the 2-mm CCI adult mice. In contrast, pediatric mice did not show a significant decrease in the distance (cm) travelled or the mean total number of errors during training. Unlike the pediatric group, the adult group exhibited a significantly higher average moving speed during training in sham rather than 2-mm CCI mice.
Retention phase: An unpaired t-test showed significant differences in the primary latency to the target hole and to adjacent holes between the pediatric and adult sham mice and between the sham and 2-mm CCI adult mice, indicating that the adult 2-mm TBI mice had memory deficits (Fig. 4E). However, the primary path length to the target hole was not significantly different between the groups (Fig. 4F).
Passive avoidance performance
Regardless of TBI, pediatric and adult mice demonstrated a good ability to learn the task as they avoided coming into the chamber in which they were given a shock on the previous day and 4 days ago. The step-through latency was not significantly different between the groups (Fig. 5).
Anxiety-like behavior
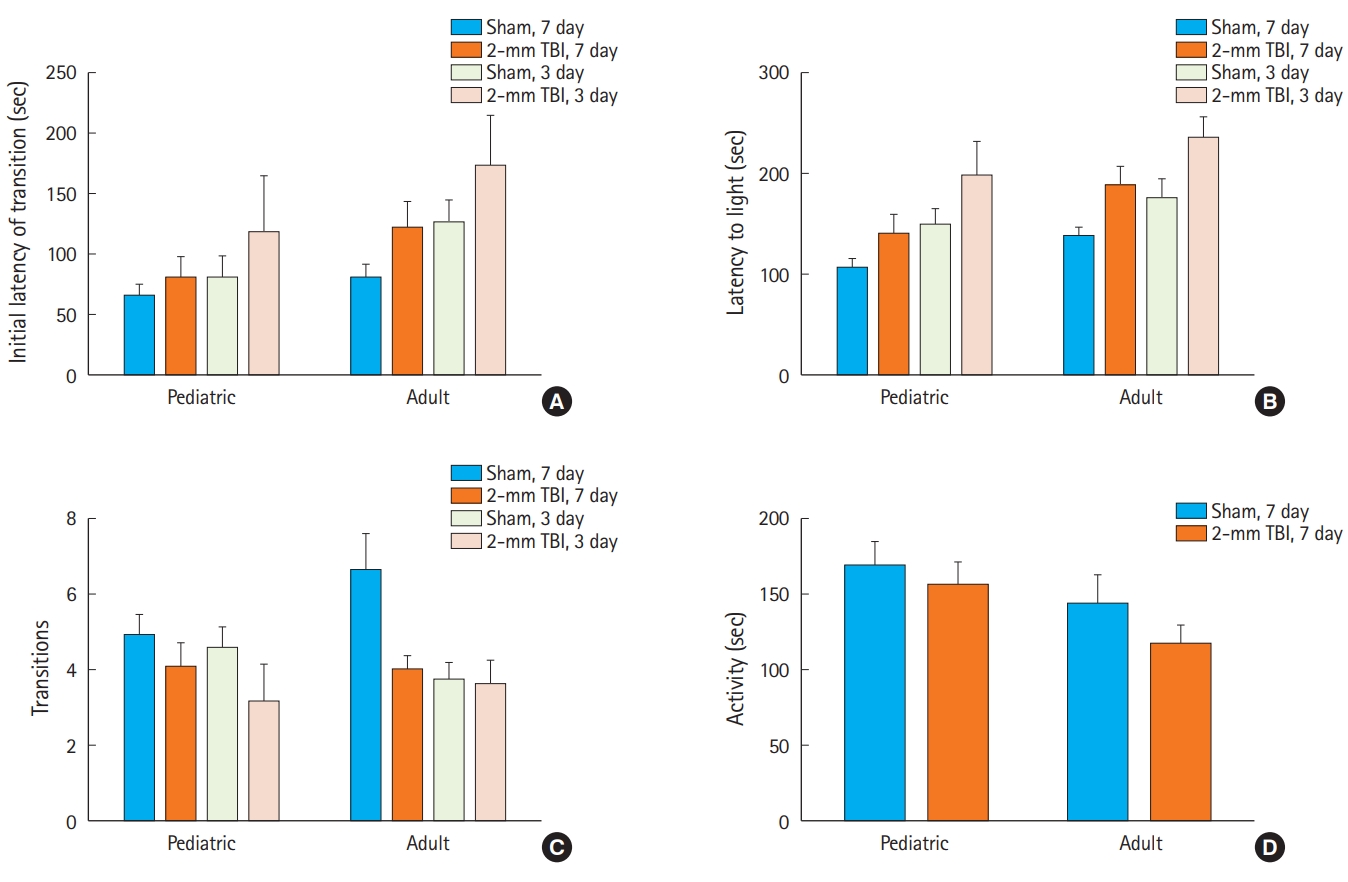
In the light-dark box test, both pediatric and adult mice after 2-mm CCI did not show a significant difference in the initial latency of transition (Fig. 7A), time spent in the light compartment (Fig. 7B), or transition times (Fig. 7C) from sham mice. Interestingly, adult mice spent significantly more time in the light compartment than pediatric mice. Notably, 2-mm CCI did not also lead to a significant reduction in immobility time relative to sham mice in both the pediatric and adult groups in the tail-suspension test (Fig. 7D).
Motor performance
There was a significant difference between the adult and pediatric 2-mm CCI at 3 days, in the performance time and falling velocity when evaluating motor performance using the accelerating rotarod (P<0.05). However, no significant differences were observed between the sham and 2-mm TBI mice in each age group for the performance time and falling velocity when using the accelerating rotarod (Fig. 8).
DISCUSSION
In addition to analyzing mortality rates and physiological variables, this study characterized histopathological responses and neurobehavioral outcomes in pediatric and adult mice following CCI. Pediatric mice had a higher mortality rate than young adult mice following CCI. Although there are similarities between head injury in the developing and adult brain, pediatric brain injury poses unique challenges [22]. The primary injury is generally more diffuse in children because of a greater head-to-torso ratio and brain-water content [22]. Clinical studies have demonstrated that the developing brain may also be more vulnerable to traumatic injury than the adult brain [22-24]. However, in our study, we applied the same impact for pediatric and adult brains.
Compared with other TBI models, such as the fluid percussion brain injury blast wave and weight drop, the CCI model is an easy and accurate experimental TBI research tool that uses a mechanical system to deliver physical impact to the exposed mouse dura [25-27]. It can precisely control many injury parameters, such as velocity, position, area, time, and depth of impact, accurately producing nearly identical injuries across groups of mice [26]. Therefore, the CCI model can overcome the epidemiological complexity and age-related inconsistent clinical findings seen in human TBIs.
In this study, the CCI was performed over the right parietal cortex and near the middle portions of the bregma, lambda, and sagittal suture. Thus, hippocampal injury, ranging from mild to severe, similar to that occurring in humans, could be observed. In a pilot experiment, after mild 1.0-mm impact in adult male mice, there appeared to be no apparent injury to the hippocampus with limited cortical lesions. However, a severe 2.0-mm impact evoked hippocampal injury with a graded pattern. Our graded injury pattern is similar to that reported by other studies [28-30]. This histopathological graded injury in the cortex and hippocampal area is closely related to differential behavioral and functional findings [31,32]. Some experimental models, such as stroke [33,34], cardiac arrest [16], and TBI [35,36] models, have suggested age-dependent differences in neuronal ischemic vulnerability. Although the mechanisms have not been fully investigated, numerous neurochemical and physiological changes at different ages could play an important role in the responsiveness to various ischemic insults, which leads to an age-dependent discrepancy in excitotoxicity, cerebral edema, oxidative stress, inflammatory response, and blood-brain barrier permeability after brain injury [35,37-39]. An important observation in the present series of histological investigations is that adult mice showed significantly more similar degree of histological injury at the acute stage compared with pediatric mice. However, there was a significant difference in the magnitude of injury volume between 3- and 7-days after adult TBI.
Although the influence of age differences on pathophysiological processes after TBI have not been completely investigated, TBI has a complex pathophysiological process consisting of several overlapping phases, including primary injury, secondary injury, and regeneration. After the primary injury, such as focal hematomas, contusions, or diffuse injury, secondary injury mechanisms are initiated that manifest over a period of hours to days through complex biochemical and physiological processes. This secondary injury could be exacerbated by the aging process [40,41].
Severe TBI presents significant cognitive and/or affective dysfunction that can have perpetual adverse consequences on the quality of life. Cognitive problems following TBI include decreased memory and learning abilities, impaired attention and concentration, reduced processing speed, word finding difficulties, and impaired executive functioning (i.e., reduced ability to plan, sequence, prioritize, think flexibly, abstract, or problem solve) [7]. The cognitive processes that result in memory impairment following TBI are not fully understood. Although experimental animal studies have demonstrated that TBI induces both retrograde and anterograde memory deficits in tests such as the Morris Water maze, it is uncertain as to what types of learning abilities or memory are affected following TBI, and which anatomical structures are responsible for the observed deficits [42]. Our study demonstrated that the Barnes circular maze was an efficient cognitive task to assess spatial/non-spatial learning following CCI injury in adult mice, but it was impossible to assess spatial learning impairment in pediatric mice.
We used the PAT to evaluate amygdala-dependent emotional memory after TBI. Passive avoidance performance is an adaptive response to a stressful experience that serves as a measure of learning and memory. This test is based on the innate preference of mice for the dark compartment of the apparatus, and suppression of this innate preference following exposure to an inescapable shock involves NMDA receptors in the amygdala and hippocampus in the formation of post-training memory [43]. However, the PAT in our study did not show significant memory impairment after 2-mm focal TBI of the right parietal lobe in both pediatric and adult mice. Khalin et al. [44] showed that with a weight-drop closed head injury, similar results were achieved in mice with TBI from both experimental groups on days 1 and 4. However, on day 7, the mean latency was significantly shorter in mice with TBI caused by 250 g of falling weight than in sham-operated mice and mice with TBI caused by 200 g of falling weight. This indicates that at 1 week after injury, mice with TBI caused by 250 g of falling weight developed cognitive deficits. Further long-term studies are warranted to clarify this observation.
In addition to cognitive changes that occur following TBI, most individuals with TBI are diagnosed with one or more newly acquired psychiatric diagnoses, such as anxiety-like behavior and depression, after injury [45].
A growing body of experimental evidence indicates that post-TBI anxiety disorders do not show a clear association with injury severity, although they appear to be more prevalent following mild TBI, unlike cognitive impairments that are worsened with an increased severity of injury [46]. Evaluation of anxiety states by using the open-field test, light/dark test, and tail suspension test in our study did not show definite differences in anxiety-like behavior between sham and 2-mm focal TBI on the right parietal lobe in both pediatric and adult mice.
The study has several limitations. First, we did not evaluate female mice in our study. Second, we did not test other strains of mice. Third, we lesioned the focal right parietal lobe; we need to further investigate other types of focal and diffuse brain injuries. Therefore, extensive testing on behavioral and electrophysiological outcomes is necessary to evaluate the sex effect in detail. Fourth, we did not administer a 2.5- or 3.0-mm impact because of its high mortality rate after surgery. Fifth, we used tiletamine/zolazepam for anesthesia, which is known to block NMDA receptors [47]. Hence, the anesthetic could affect the result of the neurobehavioral tests such as the PAT. Finally, mice were sacrificed 3 and 7 days after injury; thus, the long-term effects (more than 2 months) on histopathological outcomes were not assessed.
We demonstrated age-dependent differences in the loss of cortical mantle volume and neurobehavioral outcomes at an acute stage following CCI. There were age-dependent differences in the rate of injury progression following severe impact and spatial memory deficits in the experimental TBI models. Pediatric mice demonstrated faster injury progression than adult mice, and adult mice exhibited spatial memory deficits following CCI. Understanding these differences could be an important first step in research on the age-dependent progression of the pathogenesis and developing age appropriate therapeutics for TBI.